
Introduction
The human gastrointestinal (GI) tract holds a diverse and metabolically active microbial ecosystem that initiates colonizing during the neonatal period and is modulated throughout life by diet, environmental exposure, and host physiology. Colonization of microbes starts first along the oral cavity, where saliva (and oral mucosa) is a repository of thousands of species and millions of microorganisms that are swallowed every day. Yet, only a small number of them manage to survive and colonize the intestine due to several physiological defense barriers such as the acid environment in the stomach, bile acid (BA) secretion, digestive enzymes, and antimicrobial peptides in the duodenum and gut (1). Nonetheless, the intestinal tract harbors one of the most complex microbial ecosystems in the body and is a key determinant of human health that is increasingly being recognized as a major factor influencing a wide range of related disorders. Substantial evidence has emerged that the composition of gut microbes is a major determinant of host metabolic and immune homeostasis and disease susceptibility. Microbial diversity and functional changes (dysbiosis) have been repeatedly implicated in inflammatory bowel disease (IBD), celiac disease, irritable bowel syndrome (IBS), colorectal cancer (CRC), chronic liver disease, pancreatic diseases, and numerous metabolic and immune-mediated extraintestinal disorders (2–4). These are recent large-scale microbiome initiatives like the Human Microbiome Project and longitudinal host–microbiome association studies, confirming that microbial metabolites, immune signaling molecules, and host-derived modulators interdependently regulate systemic physiology. The gut microbiota (GM), intestinal epithelial barrier, and liver act as a connected defense network integrated for metabolic homeostasis, biotransformation pathways, and immune regulation to modulate environmental stressors (e.g., diet alterations, pollutants, and/or xenobiotic exposure) (5). Additionally, gut commensals carry out essential biological activities such as carbohydrate metabolism and vitamin biosynthesis, regulation of innate and adaptive immunity, and colonization resistance via interspecies niche for ecological nutrients in order to inhibit pathogen colonization (6). Rapid advances in nanomedicine and material science have gained considerable interest in
engineered nanoparticles (NPs) for diagnostic, targeted drug delivery, biosensing, imaging, deoxyribonucleic acid (DNA) nanotechnology, and food applications (7). Advanced in vitro models, such as intestinal structures embedded in hydrogels or microfluidic gut-on-chip systems, are now more accurately generated to mimic native human intestinal pathophysiology and thus have also transformed microbiome research through material innovation. Such insights have created a critical milestone with opportunities to define host–microbe–material interactions through these polymer scaffolds that have powered biomaterials towards their dual use as materials for studying gut communities and tools for modifying in situ GM (8). While, in parallel, the increasing use of engineered nanomaterials (ENMs) across many fields, such as industrial applications, food, and biomedicine, result in widespread exposure across the human population via food, drinking water, supplements, and therapeutics. Due to their unique nanocharacteristics, such as nanoscale dimension, tunable surface chemistry, high surface-area-to-volume ratio, and controlled biodegradability/nontoxicity properties, nanomaterials exhibit considerable biological reactivity in the GI environment (9). Nanomaterials such as metallic and metal oxide NPs [TiO2, ZnO, Ag2O] have reported important antimicrobial activity against Escherichia coli, Bacillus subtilis, and Staphylococcus aureus (10, 11), in particular the broad-spectrum toxic effect of nano-TiO2 and ZnO on both Gram-positive bacteria, Gram-negative bacteria, and some fungal strains. Although these antimicrobial properties make nanomaterials appealing for therapeutic and antimicrobial applications, the lack of microbial specificity may result in disrupting beneficial bacteria, inducing dysregulated metabolic production by beneficial bacteria, thereby losing mucosal barrier function in addition to long-term ecological imbalance in the gut. Most importantly, gut microbial composition and function are driven by a complex interplay between host molecules (microRNAs, antimicrobial peptides, mucus, immunoglobulin A, and hormones), microbial signals (lipopolysaccharides, quorum-sensing molecules, and short-chain fatty acids), and dietary factors of pharmaceuticals nanostructured particles that are ingested either intentionally or unintentionally into the GI tract (12). The microenvironment of the intestinal lumen represents a dynamic cross-talk between ENMs, mucus, epithelial cells, immune mediators, digestive enzymes, and resident microbiota. Biodegradable nanomaterials can be degraded and their physicochemical behavior, bioavailability, and biological activity changed by the action of gut enzymes or microbial metabolism. Instead, nanomaterials are able to impact microbial diversity and metabolic output as well as host immune signaling that creates systemic physiological effects. Although interactions between nanomaterials and gut microbiomes are increasingly recognized, our understanding of how the physicochemical properties of a specific nanomaterial alter microbial communities and host signaling pathways, which in turn modify therapeutic outcomes, continues to be very limited. Recent studies have mainly centered on the nanotoxicological effects, while a relatively small number of studies were dedicated to rationally designing microbiome-responsive nanomaterials that are capable of specifically modulating gut microbial composition, delivering microbiome-targeted therapeutics, restoring eubiosis, and improving therapeutic efficacy in dysbiosis-related diseases. Addressing this gap is necessary for the clinical efficacy of genomically enabled, next-generation precision nanomedicines based on targeted modification of the gut microbiome with minimal unintended disruption to host–microbial homeostasis.
The nanomaterials and gut interaction
Nanomaterials (NM)-based drug delivery systems are emerging technologies with great potential in cancer therapy due to their multifunctional properties, including targeted drug delivery, stimulus-responsive release, improved bioavailability, and multimodal therapeutic effects. There are at least three routes (inhalation, intravenous, or oral delivery routes) to administer these nanocarrier systems. Among them, oral nanomedicine is of interest, as it may be noninvasive as well as provide localized GI targeting. Yet upon oral presentation, NMs face the constantly altering GI microenvironment, and their physiochemical behavior, stability, and therapeutic function are heavily reliant on biological barriers, which include the mucus layer, intestinal lumen, epithelial barrier, and lamina propria that jointly manage NM biodistribution, retention and host interaction (13). Current research has become emphasized to provide knowledge regarding the interaction of ENMs not only with host tissues but also with the gut microbiome capable of mediating immune signaling or providing metabolites regulating intestinal homeostasis. However, results between studies are still heterogeneous and in some cases even conflicting. For example, metallic NPs Ag metal and ZnO can impact antimicrobial activities and suppression of pathogenic taxa, but multiple studies also show associated loss of beneficial commensal populations, decreased short-chain fatty acid production, compromised mucosal barrier integrity and impaired mucus production. Conversely, although polymeric, lipid-based and biodegradable nanocarriers have been generally associated with better biocompatibility and less disruption of microbiota dysbiosis; their long-term ecological effect on microbial community structure and metabolic function has not been extensively characterized. These conflicting results also highlights that microbiome responses to nanomaterials are likely less predictable and far more dependent on the specific characteristics of each combination of composition, size, surface chemistry, dose, exposure duration and host physiological state than anticipated. Notably, nanomaterials interact with gut microbiome in both directions. Nanomaterials can change the composition and functional gene expression as well as metabolic outputs of microbes in parallel with gut microbial enzymes and metabolites transforming nanomaterial physicochemical characteristics through biodegradation, biotransformation, surface modification, or aggregation, resulting in changes to bioavailability, residence time, and therapeutic efficacy of encapsulated drug delivery NPs (14). Although a number of publications have documented this bidirectional interplay between nanomaterials and microbes, several practical limitations hamper our current understanding. Moreover, as highlighted in this review, the majority of studies published to date have mainly focused on compositional microbial changes but not functional outcomes (i.e., metabolite production, immune modulation, and/or host signaling cascades). Together, these limitations underscore an urgent requirement for systematic mechanistic studies that incorporate nanomaterial design parameters, microbiome functional profiling, and the host physiological responses. This knowledge will be critical for the different types of nanotherapeutics targeting the microbiome to enhance their therapeutic effect while minimizing disruptions in gut microbial ecology and host homeostasis.
Implications of nanodrug formulations for anticancer drugs
The complex involvement of GM in the regulation and metabolism of anticancer drugs was recently confirmed by several studies. In addition to host metabolism, GM-mediated enzymatic transformation of chemotherapeutic drugs can affect therapeutic potential and structural stability. In particular, common anticancer drugs that are widely used, such as five-fluorouracil, irinotecan, and cyclophosphamide (the conversion of these drugs is mainly dependent on the GI tract microbiota) generate small metabolite products like short chain fatty acids (SCAFs), including butyrate BA derivatives (15–17). These microbial conversions may enhance or diminish drug action, suggesting that the crosstalk between host therapeutic and microorganism activity needs to be viewed bidirectionally. Although these nanodrug technologies promise improvement of the new drug candidate, their application at this interface presents advancing challenges and emerging possibilities. Nanodrug formulations can enhance drug solubility and permeability, which leads to a better therapeutic effect and lower systemic toxicity effects compared with small molecular drugs. Additionally, technologies for surface modification and nanostructure engineering have been developed so that these systems can be held in the GI tract longer, allowing for increased exposure to GM. Furthermore, extended contact time might further increase the effectiveness of biotransformation processes mediated by microbes where colonized microbial flora are thought to be an important factor in determining effectiveness and fate during nanodrug-based antitumor therapies (18).
Alterations in gut microbiota and dysbiosis
The GM is involved in host defense against pathogens via processes such as colonization resistance and immune regulation, including production of metabolites (19), most notably SCFAs, vitamins, and BA derivatives. When the gut microbiome homeostasis is disturbed because of various factors, it will result in an imbalance status defined as dysbiosis (19). Dysbiosis results in less diversity, which may be reduced microbial diversity or alterations of taxon composition. There are no specific profiles of dysbiosis, but healthy subjects share certain common compositional patterns. According to the Anna Karenina principle, a multitude of compositional and functional defects may variably disturb each of these dysbiotic states (20–22). The GM is a complex ecosystem with thousands of microbial strains and species, represented in many ways between individuals. Microbial diversity, metabolic capabilities, functional cooperation, and interspecies signaling are critical components of maintaining intestinal and systemic homeostasis (23). Dysbiosis could mediate disease by altering functional pathways of microbial metabolism or gut barrier permeability or through modulation of immune regulatory pathways (24). Therefore, a great focus is being put on controlling the microbial balance for health and disease prevention. Diet is the primary driver of gut microbiome composition. Metabolic cross-talk among dietary fiber and gut microbes is important for maintaining the gut ecosystem (25).
Interaction between biological systems and nanomaterials
Nanotechnology is a significant area of science that has dramatically impacted many fields, including solid-state physics, materials sciences, chemistry, biology, computing science, and engineering. Its use for biomedical research applications has increased rapidly over the past few years, with novel applications particularly in oncology and cancer therapy (26) (Figure 1). Because of their unusual physicochemical and biological characteristics, NMs have been well studied as platforms for drug and gene delivery, diagnostic biosensing, and therapeutic monitoring. Many classes of nanosystems exist, including metallic NPs (gold (Au), silver (Ag), copper (Cu), and zinc (Zn) NPs), which have been developed over the past 20 years due to their specific catalytic, chemical, and optical properties. The predominant biomedical applications of their types are controlled drug release of anticancer drugs and targeted therapy (27).
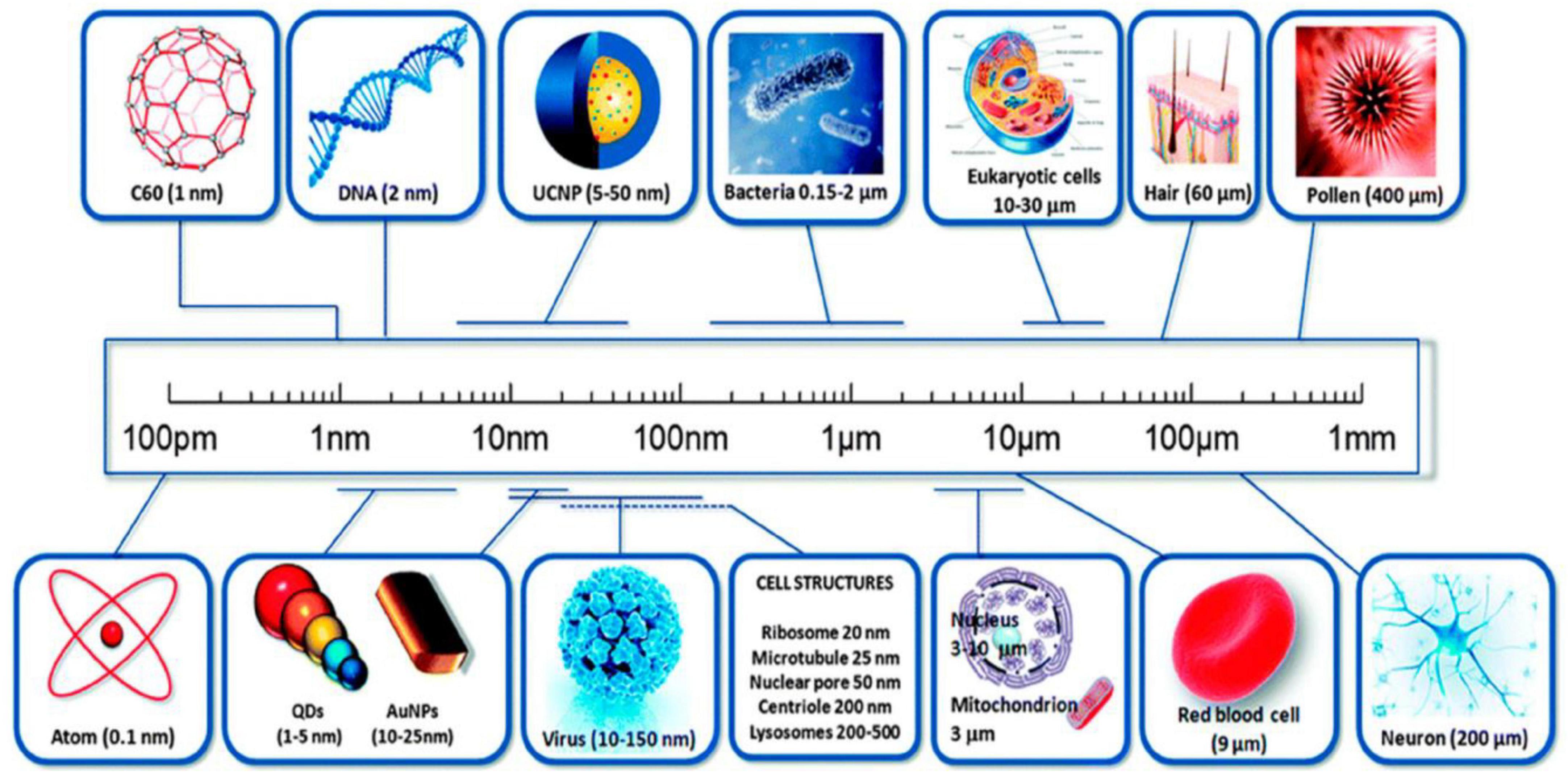
Figure 1. A comparison of nanomaterial particle sizes. (Image adopted from ref. (26)).
The majority of the nanomaterial synthesis is based on either bottom-up or top-down methods.
In a top-down approach, bulk material is grinded into nanoscale through complex industrial method, such as precision engineering. Nanotechnologies also play an important role in precision engineering, the foundation of the microelectronics industry, wherein cutting tools at nanoscale, such as diamond or cubic boron nitride, high-resolution sensors for measurement and feedback systems, and advanced servo-drive and numerical control are used. Surface structuring is done by utilizing photons or ions or electrons followed by material depositions on the pre-patterned surfaces for building nanoscale materials. On the contrary, the bottom-up strategy assembles nanostructures from atomic or molecular building blocks through controlled physicochemical events in nanoscale dimensions (1–100 nm). Chemical synthesis provides simple molecules that can be applied directly or reorganized into advanced composites. This can be done via self-assembly by having the atoms or molecules arrange themselves to form ordered structures through physiological interactions that they are thermodynamically capable of engaging in or positional assembly of the minimum limit to bottom-up fabrication whereby a grid enclosing single atoms or clusters is seeded with their positions predefined (28) (Figure 2).
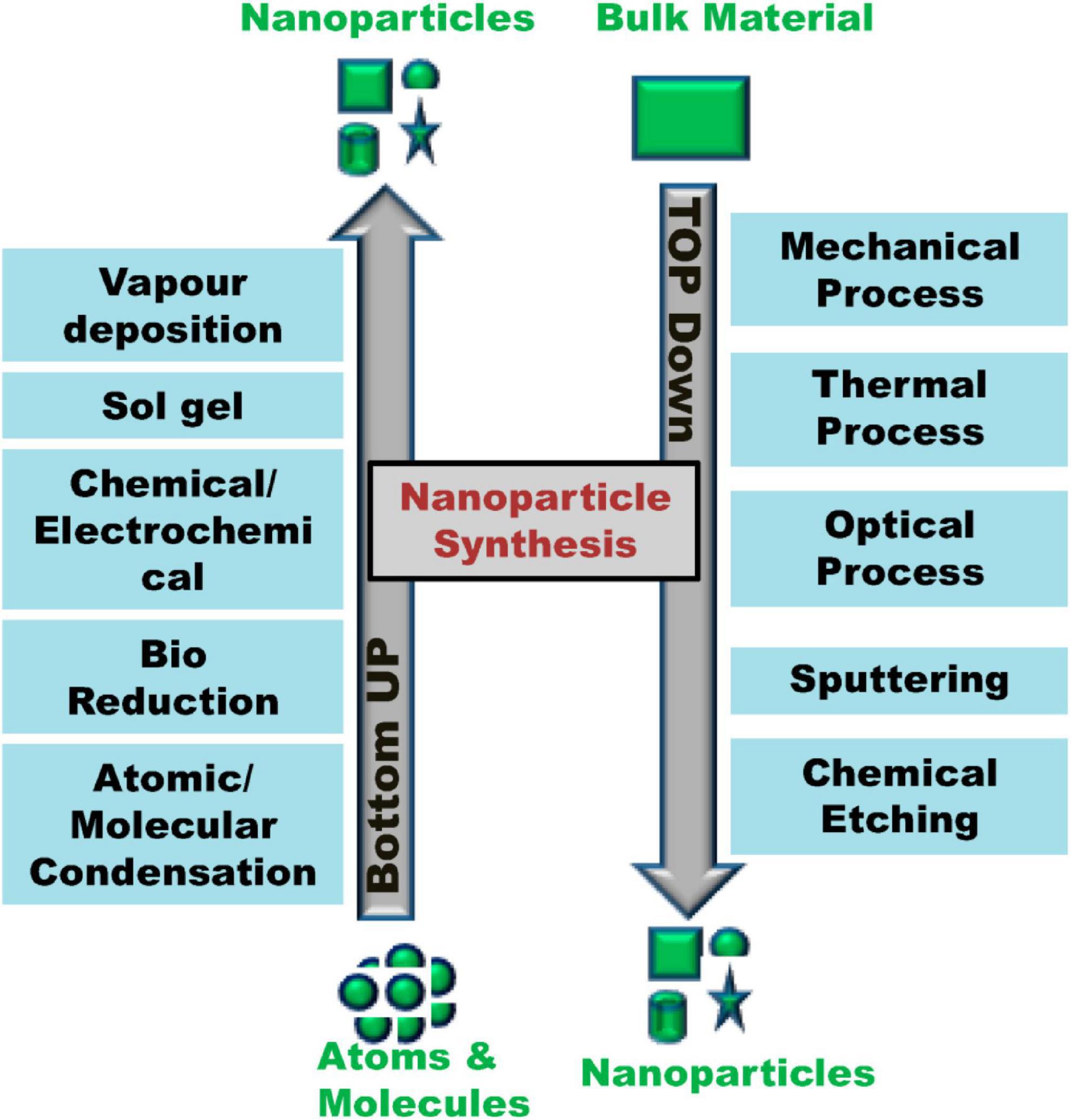
Figure 2. Top-down and bottom-up approaches for nanoparticle synthesis. (Image adopted from ref. (26)).
Nanoparticle synthesis, particularly in the context of drugs, has transformed the drug production landscape. Currently, modern engineering nanodrug systems can enhance the pharmacological safety profile by improving drug solubility, stability, and bioavailability and at the same time minimizing systemic toxicity through site-specific delivery. Van de Water and coauthors reported that efficient drug delivery systems came from an extensive optimization process with low predictability. With respect to data-driven design and underlying mechanisms, existing nanotechnology promotes a more systematic and efficient development pathway. In turn, personalized medicine has transformed the nanocarrier system design towards oriented approaches based on some patients’ specific features more involved in achieving higher therapeutic efficacy and clinical benefit (29). Nanotechnology in today’s world has become a commonly used tool among various aspects of biomedical and environmental perspectives. Besides thoroughly reviewing therapeutic NPs and biosensors, some have emerged as potential diagnostic agents for nanomedicine or monitoring tools for environmental purposes and effective biological sensing platforms. Progress has also been achieved in the development of optical plasmonic sensors based on nanomaterials that exhibit ultrahigh sensitivity, high-efficiency nanoparticle detection, low cost, and biocompatibility advantages, as well as remote analysis (30). Generally, nanotechnology continues to evolve since its initial development into technology use.
Biocompatibility, toxicity, and biodistribution of nanomaterials in the GI tract
The European Society for Biomaterials (1986) said that “biocompatibility is the ability of a material to perform with an appropriate host response in a specific application” (31). Various nanostructured materials designed particularly with carbon, silica, and certain metals have been studied for biomedical application, having distinctive properties based on the morphology, such as spheres, tubes, and rods (32). As soon as NMs penetrate the gastro intestinal tract (GIT), they encounter a diverse and complex intestinal environment characterized by a mucus-rich mucosa system. The elimination of NMs from the GI lumen is mediated by the excretion via feces driven by mucous turnover and intestinal motility, thus potentially leaving an incomplete proportion of the dose ratio, enabling the presence of NMs in the gut lumen after follicle-associated epithelium nano material (FAENM) exposure that may modulate GM composition and function as well as disrupt integrity (33). GM also shapes this interface by regulating mucin production. Commensal and pathogenic bacteria, including Lactobacillus rhamnosus and E. coli, have been shown to induce the production of specific mucins, pointing that there is an ongoing communication between bacterial activity and commensal mucus barrier functions (34). Initial progression of NM interaction in the intestine begins with its adsorption on protective layers within the extracellular matrix, followed by their passage to reach the surface of epithelial cells. SCFAs, which represent metabolites of the GM, are relevant immune-modulating factors at both mucosal and systemic levels; therefore, NM-mediated depletion of SCFA-producing bacteria may disrupt the homeostasis of the immune system and promote inflammation (35). NPs can be taken up by cells and processed in different intracellular compartments, resulting in the modulation of cellular function defined as beneficial or unfavorable. Physiological studies have indicated that several NPs increase production of reactive oxygen species (ROS) and damage organelles, membranes, nuclei, and enzymes. These changes lead to a reduction in both adenosine triphosphate (ATP) synthesis and protein expression disorders stimulating apoptotic pathways (36). Biomaterials are typically viewed as foreign bodies that may cause inflammation, and size is a common mediator of nanoparticle-induced inflammation. Example: Below 20 nm polyacrylic acid–coated gold nanoparticles (AuNPs) were shown to promote fibrinogen-mediated activation of Mac-1 through the receptor on monocytes, resulting in NF-κB signaling and cytokine production. On the other hand, larger particles (≥20 nm) are unable to mediate this receptor activation equivalent, indicating that a <20 nm particle size is crucial in response to inflammatory events (37). In pharmacokinetics studies aiming at drug metabolism and immune modulation, the crosstalk between GM and anticancer agents was highly complex (13). Consistent with these findings, complementary omics-based studies have demonstrated that metastable exposure conditions for NMs can affect bacterial physiology by modulating specific cellular processes such as membrane transport and subsequent signal transduction, energy metabolism, and also quorum sensing and biofilm formation pathways. These networks can be affected when incorrect regulation is implicated, which may change the gut microbial composition (38).
Nanoparticle–microbe crosstalk: mechanistic insights
In the last 10 years, numerous research studies have focused on the application of NPs in humans and animals. Progress in the synthesis and functionalization of metallic, magnetic, and polymeric NPs; liposomes; nanoemulsions; and nanogels has greatly enhanced their effectiveness against drug-resistant bacteria, fungi, and viruses. These nanoplatforms not only secure better delivery and stability of antimicrobial compounds but also have been shown to inhibit quorum sensing and disturb biofilm architecture as well as enhance the therapeutic efficacy of conventional antibiotics (39). These advances have made nanotechnology a prospective approach for combating antimicrobial resistance, especially in cases where there is low penetration, rapid degradation, or poor bioavailability of conventional antimicrobial agents. That means for NM-microbe interactions come into play due to the high structural diversity of microorganisms and highly dynamic physicochemical conditions in complex biological systems like the GI tract, host-associated microbiomes, and ecological microbial communities (40). Many studies are focused on NP-enabled antimicrobial therapy, but it should be emphasized that naturally occurring NP-microbe interactions can also play a key role in disease progression, microbial ecology, and host physiology (41). Although research interest is rising, the field is still at an early stage, and the physicochemical principles underlying selective nanoparticle–microorganism interactions are yet incompletely defined in terms of microbial susceptibility that is microbe- and host-dependent (42). Direct interactions of NPs with biological membranes are one of the main mechanisms underlying their antimicrobial activity. Membrane phospholipids of many bacterial cells and surface-exposed polysaccharides such as lipopolysaccharide contribute a negative charge, which in turn facilitates electrostatic attraction between positively charged NPs or surface-functionalized NPs. For certain nanosystems, they can pass through the cell membrane and localize intracellularly, interacting with ribosomes and DNA as well as intracellular enzymes or membrane-bound respiratory proteins to disrupt replication and transcription of genes followed by inhibition of ATP production. Similarly, the antimicrobial mechanism is induced through the formation of ROS (43). NPs with metallic and metal oxide characteristics such as AgNPs, ZnO, CuO, TiO2, and iron oxide nanoparticles (FeNPs) have been reported to produce ROS under physiological or photoactivated conditions; catalytic production of ROS occurred at low concentrations. This in turn results in the production of hydroxyl radicals, superoxide anions, singlet oxygen, and hydrogen peroxide, thus creating oxidative stress, which then surpasses microbial antioxidant defense capacity. Increased amounts of ROS cause lipid peroxidation of cellular membranes, oxidation of both enzymes and proteins, DNA strand breaks, mitochondrial dysfunction in fungi, and irreversible damage to microbial macromolecules. This oxidative damage promotes both bactericidal and fungicidal activity as well as adaptive stress response pathways in microbes (44). Conversely, marginal nanoparticle exposure can stimulate adaptive metabolic remodeling that facilitates microbial stress resistance or enrichment of resistant populations, suggesting that nanoparticle impacts do not always show linear inhibition. For example, broad-spectrum antimicrobial NPs may inhibit favorable commensals like Lactobacillus, Bifidobacterium, and butyrate-producing bacteria while enhancing a population of stress-resistant microbes capable of surviving oxidative stress. These ecological changes can lead to dysbiosis, modified profiles of metabolites, and an imbalance between hosts has a high-risk status for inflammation or pathogen colonization. NPs also have some key antifungal properties. Nonetheless, AgNPs exert relatively less evidence of their antifungal mechanisms compared to antibacterial ones, and the activity against pathogenic yeasts such as Candida spp. was determined to be quite similar to ionic silver (Ag+) at equally efficacious concentrations (45, 46). Some of the proposed antifungal mechanisms comprise membrane destabilization, mitochondrial ROS induction, ergosterol biosynthesis inhibition, disruption of fungal cell wall integrity, and interference with fungal biofilm formation. Developed multidimensional mechanisms render ENMs as potent agents for treating fungal infections, especially in immunocompromised cases, wherein the emergence of antifungal resistance presents a significant clinical challenge (47). Details of these interactions, including mediated mechanisms such as oxidative stress pathways, metabolic reprogramming, and community-level microbial responses, will be crucial for the design of safer and more selective nanotechnologies that promote therapeutic benefit while minimizing interference with volatile microbial ecosystems.
Therapeutic potential and clinical translational applicability of nanomaterial-based approaches for restoring gut dysbiosis
The nanoparticle-microbe interface is essential for the development of effective antimicrobial intervention. Antimicrobial effects of NPs towards bacteria and fungi could be exerted by means of different mechanisms, including generation of ROS, membrane pore formation causing disruption, cell wall damage, DNA induction and degradation, as well as disturbance in the cellular cycle (48). Pulmonary bacterial infections remain a predominant use for oral and intravenous antibiotics. Although IV administration bypasses metabolism and increases bioavailability, it does this with high toxicity due to low pulmonary targeting. Pulmonary inhalation therapy is a promising alternative that can provide targeting of the drug delivery and deposition directly in the lung and therefore decrease systemic exposure and rapid onset of action compared to the intravenous administration route (49). Among these approaches, nanocarriers are a novel technology that has high potential in effectively delivering different types of therapeutic agents (including microbial derivatives, metabolites, and nucleic acids) aimed against chronic liver diseases. Nonviral delivery vectors based on lipid NPs are very promising candidates to modulate this axis due to their better stability, target delivery, controlled release, and ability to cross biological membranes (50). NPs of various types have been shown to impact the gut microflora composition and activity. The overall gut microbiome response, such as the one to titanium dioxide nanoparticles (TiO2 NPs), is mostly a low-level host effect because of high microbial populations compared with diversity at various time points for acute conditions. TiO2 NPs also affect Lactobacillus, Firmicutes, and Proteobacteria (51, 52). Li et al. Their studies also demonstrated that TiO2 NPs accumulated in multiple tissues, including spleen, lung, and kidney; altered intestinal epithelial cell structure; and consequently led to gradual changes of GM (53). The silver nanoparticles (Ag NPs) have been reported as efficient antibacterial agents against sensitive and resistant bacteria, including GM of a broad range of species. Notably, Ag NPs reportedly increased the abundance of gram-negative bacteria and had a relatively stronger non-target effect on L. rhamnosus and E. coli (54). Considering zinc oxide nanoparticles (ZnO NPs), the antimicrobial effect would be associated with oxidative stress, physical interaction between bacteria, and oxidative assault triggered by liberated Zn ions (55). The GM is dynamic and responsive to dietary and environmental exposures. Data available on members of one of the most abundant phyla in the gut, Firmicutes, suggest that these are extremely sensitive to NP exposure. Certain genera, such as Lactobacillus, are sensitive to a range of NPs; although, Bifidobacterium directly interfaces with Ag NPs and Cu NPs but is negatively regulated by TiO2 (9). Polymeric or organic nanomaterials are mainly used as therapeutic agents in microbiota-targeting interventions. These most recently developed polymeric nanoparticle systems are made up of biodegradable polymers such as poly lactic-co-glycolic acid (PLGA) and chitosan (CS) allow targeted drug release at physiological sites through modulation of stimuli-responsive characteristics ideally suited for clinical application, including pH or enzyme expression. PLGA NPs have been reported to deliver anti-inflammatory drugs (56, 57). Chitosan nanoparticles (CNN) possess muco-adhesive properties favoring their oral delivery as carriers of microbiome-modulating agents (58). Nanomaterials can modulate multiple pathways (both metabolic and genetic) in bacteria. The metabolism of carbohydrate, lipid, amino acid, nucleotide, vitamin, xenobiotic, cofactor, and energy is altered by exposure. In addition, NPs could also interfere in the molecular mechanisms related to genetic expression mechanisms (e.g., transcription and translation, protein folding, DNA replication, and repair). Other cellular processes, such as membrane transport, signal transduction, and motility, are potentially affected as well (59) (Figure 3).
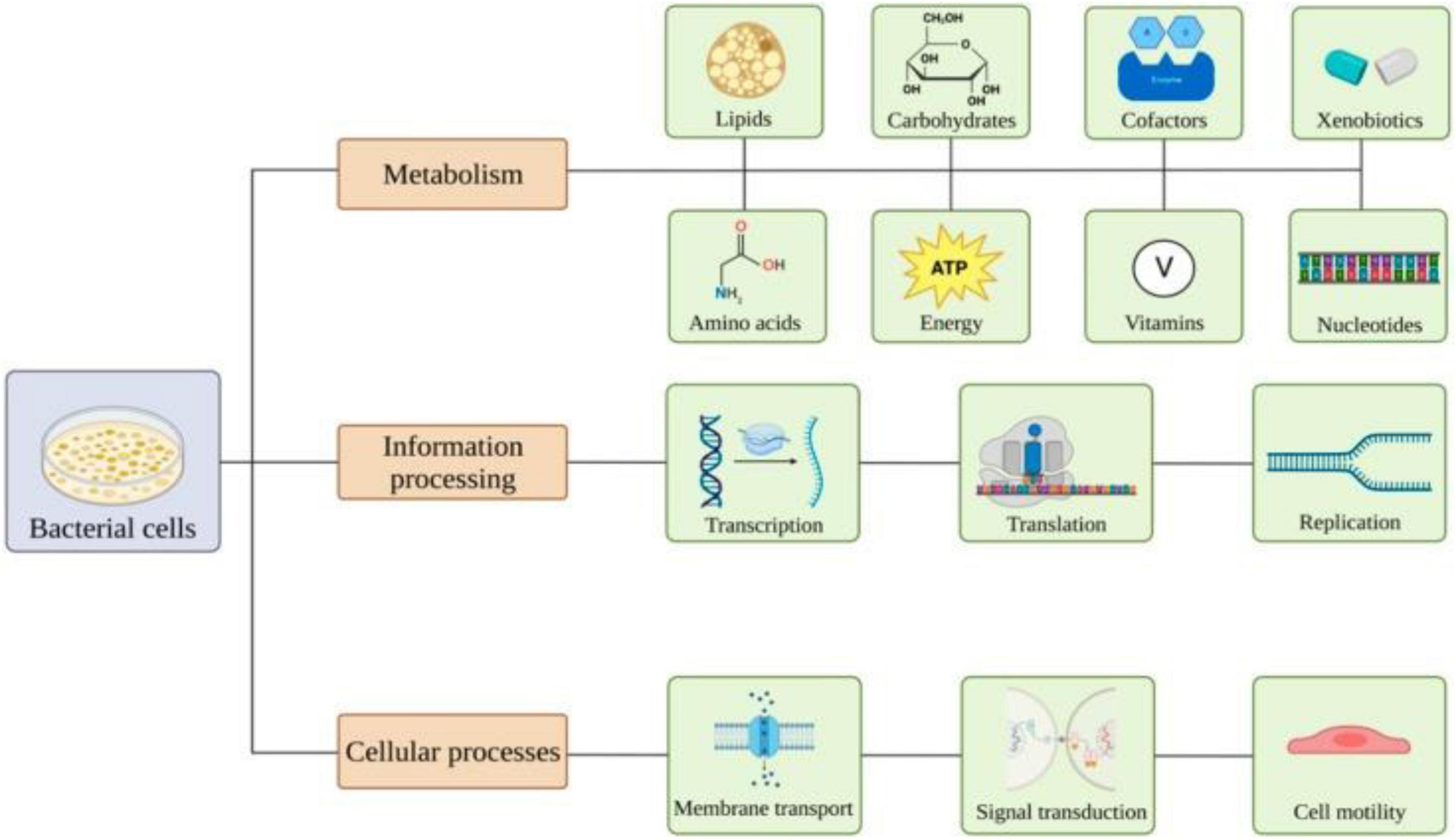
Figure 3. Overview of the effects of nanomaterials on bacterial pathways. (Image adopted from ref. (59)).
Graphene-based nanomaterials, in particular, have recently attracted interest owing to their high biocompatibility and low toxicity as compared to other nanomaterials. More and more evidence is accumulating supporting their potential use as targeted treatments, also for rare disease (60). Natural Polymeric Materials Natural polymers are renewable and eco-friendly, and they can be isolated from different biological sources, and most of them rapidly vanish into water, carbon dioxide, or other minor inorganic molecules. Their large number of functional groups in turn renders them readily chemically, physically, and nanotechnologically manipulated to produce novel hybrid materials with enhanced biomedical applications (61). Despite vaccination being considered one of the most effective medical interventions in human history, vaccines are administered through either parenteral or mucosal routes, which is performed primarily because no efficient delivery platform has been developed to direct relevant immunogenicity towards appropriate induction sites while enhancing immune responses (62). These challenges can be addressed through advancements in smart nanoparticle drug delivery like the ones proposed. The main strategies therein include performing surface engineering of the NP to ensure superior site-specific localization, selecting appropriate carrier materials for NP formulation with controlled release characteristics, and further improvisation and tailoring in nanoparticle design, ultimately aiming at enhancing targeting modulation (63). In addition to systemic function, NMs also carry potential advantages as delivery vehicles upon being administered into the GI tract. NPs can increase the stability of drugs against a highly acidic gastric environment and protect nucleic acids, peptides, probiotics, and vaccines from catalytic degradation. Therefore, they improve the bioavailability and integrity of these therapeutic agents. The residence time in GI can be extended with surface-coated or mucoadhesive NPs that are largely obtained from CS or other natural polymers (64). Importantly, NMs can be designed to selectively target the GM thereby enabling the delivery of microbial metabolites, bacteriophages, and prebiotics for microbiome regulation. This approach is beneficial in dysbiosis-related disorders, such as IBD, antibiotic-induced imbalance, and metabolic pathologies that aid in the restoration of eubiosis (65). It is the combined unique physiological characteristics of NMs consisting of controllable pore size/volume gradients, tumor-specific degradability, tunable charge plus size, and a large surface volume ratio that potentially make them a viable vehicle for GI tract-targeted therapies. The capacity of pre- and probiotics to prevent the loss of therapeutic payloads and key enhanced mucosal penetration properties through modulation of GM with the concomitant activation of mucosal integrity highlights their unique role as next-generation strategies for improving gut health and achieving a balance in the microbiome.
Clinical translation and regulatory considerations
Bioactive ions released by metallic NPs (e.g., silver, zinc oxide, titanium dioxide) might enter circulation and lead to toxicological effects outside the intestine. Importantly, frequent sublethal exposure to NPs might as well exert selective pressure on microorganisms, finally leading to nanoparticle tolerance as well as cross-resistance to antibiotics or significant alteration in the metabolic rewiring of microorganisms under natural selection, a line that is still poorly explored (66). Furthermore, for the clinical and regulatory advancement of nanomedicine, standardization frameworks for nanoparticle characterization, microbiome safety evaluation, and long-term ecological risk assessment are still lacking. Conventional pharmaceutical evaluation pipelines can be limiting for nanotherapeutics due to the strong dependence of nanoparticle efficacy and safety on numerous physicochemical attributes, including particle size distribution, zeta potential, surface chemistry, shape, colloidal stability, and batch-to-batch reproducibility. Regulatory agencies are progressively requiring complete assessment on physicochemical characterization, pharmacokinetic and biodistribution studies, immunotoxicity evaluation, analysis of potential microbiome impact, and environmental fate before clinical approval (67, 68). Although international guidelines on nanomicrobiome interactions have been harmonized, there are still no globally accepted and standardized testing protocols for the evaluation of nanopotential effects. Furthermore, scaling up can further complicate translational barriers such as reproducible synthesis, sterilization, formulation stability, storage conditions, and cost-effective scalable production under GMP. Precision-by-design strategies that emphasize material engineering combined with microbiome science, toxicology, systems biology, and regulatory science are needed for rapid clinical translation of future nanotherapeutics (69). In vivo models such as gut organoids, microbiome-humanized animal models, and gut-on-chip platforms will be critical for predicting human-relevant responses. Concurrently, regulations need to become microbiome-aware in guiding nanomedicine evaluation according to nanoparticle host toxicity and likely ecological compatibility with beneficial microbes at the site of drug action (pharmacological efficacy). Overcoming these translational and regulatory hurdles will be a critical step in the safe utilization of nanotechnology as an innovative therapeutic platform for microbial intervention, microbiome-engineering applications, and GI disease-targeted therapies (70).
Conclusion
Moreover, the growing synthesis and commercialization of ENMs have led to increasingly large use in many industrial, medical, and consumer products, which necessitate further investigation to elucidate their complex interactions with the GI tract and the GM (71). As more evidence expands, ENMs have been shown to influence gut health through their effects on microbial diversity and functional capacity, metabolite production as well as of maintaining mucosal barrier integrity and host immune defense mechanisms. Understanding the bidirectional crosstalk between nanomaterials and the gut microbiome enables both beneficial as well as deleterious systemic effects attributable to ENM exposure (72). Finally and most importantly, the nanomicrobiome interface is a fascinating new area that shows great potential as a novel platform for safer and more effective therapeutic strategies, including targeted drug delivery systems, microbiome modulation, and precision medicine approaches. At the same time, such interactions could give rise to poorly understood immunological and toxicological side effects (73). These findings are important to address the existing gaps in the nanotoxicology field but also essential to understanding the mechanisms involved in how nanomaterials induce microbiome changes, which are also crucial to achieving a better safety assessment of ENMs. The identification of early microbiome-derived biomarkers to diagnose and monitor ENM-induced dysbiosis is an exciting new path that could enable operationally effective intervention before potential long-term risks occur. Furthermore, approaches that restore the microbiome, such as probiotics, prebiotics, postbiotics, and other methods to support a healthy gut flora, may be innovative yet simple ways to mitigate the adverse effects of nanomaterials. The development and standardized guidelines for toxicological evaluation frameworks and high-end multi-omics approaches utilized in mechanistic studies, utilizing the established ENM–microbiome interaction framework, are described in this review. In conclusion, synergies between nanotechnology and microbiome science can redefine a new era of microbiome-based targeted therapeutics for more personalized medicine while supporting novel solutions to maintain human health with retention of gut microbial homeostasis.
Funding
The authors declared that financial support was not received for this work and/or its publication.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. De Vos WM, Tilg H, Van Hul M, Cani PD. Gut microbiome and health: mechanistic insights. Gut. (2022) 71(5):1020–32.
2. Van Baar AC, Nieuwdorp M, Holleman F, Soeters MR, Groen AK, Bergman JJ. The duodenum harbors a broad untapped therapeutic potential. Gastroenterology. (2018) 154(4):773–7.
3. Trebicka J, Macnaughtan J, Schnabl B, Shawcross DL, Bajaj JS. The microbiota in cirrhosis and its role in hepatic decompensation. J Hepatol. (2021) 75:S67–81.
4. Adolph TE, Mayr L, Grabherr F, Schwärzler J, Tilg H. Pancreas–microbiota cross talk in health and disease. Ann Rev Nutr. (2019) 39(1):249–66.
5. Van Hul M, Cani PD, Petitfils C, De Vos WM, Tilg H, El-Omar EM. What defines a healthy gut microbiome? Gut. (2024) 73(11):1893–908.
6. Falony G, Joossens M, Vieira-Silva S, Wang J, Darzi Y, Faust K , et al. Population-level analysis of gut microbiome variation. Science. (2016) 352(6285):560–4.
7. Forgham H, Chang Y, Wang Y, Zhu J, Liu L, Biggs H , et al. The evolution of nanomedicine: the rise of next-generation nanomaterials in cancer nanomedicine. Sci Adv. (2025) 11(43):eadx1576.
8. Yang L, Hung LY, Zhu Y, Ding S, Margolis KG, Leong KW. Material engineering in gut microbiome and human health. Research (Wash DC). (2022) 2022:9804014.
9. Ma Y, Zhang J, Yu N, Shi J, Zhang Y, Chen Z , et al. Effect of nanomaterials on gut microbiota. Toxics. (2023) 11(4):384.
10. Hajipour MJ, Fromm KM, Ashkarran AA, de Aberasturi DJ, de Larramendi IR, Rojo T , et al. Antibacterial properties of nanoparticles. Trends Biotechnol. (2012) 30(10):499–511.
11. Daou I, Moukrad N, Zegaoui O, Rhazi Filali F. Antimicrobial activity of ZnO-TiO2 nanomaterials synthesized from three different precursors of ZnO: influence of ZnO/TiO2 weight ratio. Water Sci Technol. (2018) 77(5):1238–49.
12. Lazar V, Holban AM, Curutiu C, Ditu LM. Modulation of gut microbiota by essential oils and inorganic nanoparticles: Impact in nutrition and health. Front Nutr. (2022) 9:920413.
13. Li X, Wei H, Qi J, Ma K, Luo Y, Weng L. Interactions of nanomaterials with gut microbiota and their applications in cancer therapy. Sensors. (2023) 23(9):4428.
14. Niu B, Zhang G. Effects of different nanoparticles on microbes. Microorganisms. (2023) 11(3):542.
15. Zhang F, He F, Li L, Guo L, Zhang B, Yu S , et al. Bioavailability based on the gut microbiota: a new perspective. Microbiol Mol Biol Rev. (2020) 84(2):10–1128.
16. Pant A, Maiti TK, Mahajan D, Das B. Human gut microbiota and drug metabolism. Microb Ecol. (2023) 86(1):97–111.
17. Xie Y, Hu F, Xiang D, Lu H, Li W, Zhao A , et al. The metabolic effect of gut microbiota on drugs. Drug Metab Rev. (2020) 52(1):139–56.
18. Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell. (2012) 148(6):1258–70.
19. Bidell MR, Hobbs AL, Lodise TP. Gut microbiome health and dysbiosis: a clinical primer. Pharmacotherapy. (2022) 42(11):849–57.
21. Verma S, Dutta SK, Firnberg E, Phillips L, Vinayek R, Nair PP. Identification and engraftment of new bacterial strains by shotgun metagenomic sequence analysis in patients with recurrent Clostridioides difficile infection before and after fecal microbiota transplantation and in healthy human subjects. PLoS One. (2021) 16(7):e0251590.
22. Ke S, Pollock NR, Wang XW, Chen X, Daugherty K, Lin Q , et al. Integrating gut microbiome and host immune markers to understand the pathogenesis of Clostridioides difficile infection. Gut Microb. (2021) 13(1):1935186.
23. Shen Y, Fan N, Ma SX, Cheng X, Yang X, Wang G. Gut microbiota dysbiosis: pathogenesis, diseases, prevention, and therapy. MedComm. (2025) 6(5):e70168.
24. Zhang Y, Wang H, Sang Y, Liu M, Wang Q, Yang H , et al. Gut microbiota in health and disease: advances and future prospects. MedComm. (2024) 5(12):e70012.
25. Sonnenburg ED, Smits SA, Tikhonov M, Higginbottom SK, Wingreen NS, Sonnenburg JL. Diet-induced extinctions in the gut microbiota compound over generations. Nature. (2016) 529(7585):212–5.
26. Bayda S, Adeel M, Tuccinardi T, Cordani M, Rizzolio F. The history of nanoscience and nanotechnology: from chemical–physical applications to nanomedicine. Molecules. (2019) 25(1):112.
27. Gnach A, Lipinski T, Bednarkiewicz A, Rybka J, Capobianco JA. Upconverting nanoparticles: assessing the toxicity. Chem Soc Rev. (2015) 44(6):1561–84.
28. Iqbal P, Preece JA, Mendes PM. Nanotechnology: the “top-down” and “bottom-up” approaches. In: Gale PA, Steed JW editors. Supramolecular Chemistry: From Molecules to Nanomaterials. Wiley. (2012).
29. Jadhav V, Roy A, Kaur K, Roy A, Sharma K, Verma R , et al. Current advancements in functional nanomaterials for drug delivery systems. Nano Struct Nano Objects. (2024) 38:101177.
30. Uniyal S, Choudhary K, Sachdev S, Kumar S. Nano-bio fusion: advancing biomedical applications and biosensing with functional nanomaterials. Optics Laser Technol. (2024) 168:109938.
31. Duncan R, Izzo L. Dendrimer biocompatibility and toxicity. Adv Drug Deliv Rev. (2005) 57(15):2215–37.
32. Ketabchi N, Naghibzadeh M, Adabi M, Esnaashari SS, Faridi-Majidi R. Preparation and optimization of chitosan/polyethylene oxide nanofiber diameter using artificial neural networks. Neural Comput Appl. (2017) 28(11):3131–43.
33. Qiao X, Bao L, Liu G, Cui X. Nanomaterial journey in the gut: from intestinal mucosal interaction to systemic transport. Nanoscale. (2024) 16(41):19207–20.
34. Limage R, Tako E, Kolba N, Guo Z, García-Rodríguez A, Marques CN , et al. TiO2 nanoparticles and commensal bacteria alter mucus layer thickness and composition in a gastrointestinal tract model. Small. (2020) 16(21):2000601.
35. Tang M, Li S, Wei L, Hou Z, Qu J, Li L. Do engineered nanomaterials affect immune responses by interacting with gut microbiota? Front Immunol. (2021) 12:684605.
36. Peng C, Lu W, Fang Y. An insight into the effect of food nanoparticles on the metabolism of intestinal cells. Curr Opin Food Sci. (2022) 43:174–82.
37. Aljabali AA, Obeid MA, Bashatwah RM, Serrano-Aroca Á, Mishra V, Mishra Y , et al. Nanomaterials and their impact on the immune system. Int J Mol Sci. (2023) 24(3):2008.
38. Zhang Y, Mortimer M, Guo LH. Interplay between engineered nanomaterials and microbiota. Environ Sci Nano. (2020) 7(9):2454–85.
39. Kumari M, Klodzinska SN, Chifiriuc MC. Microbe-nanoparticle interactions: a mechanistic approach. Front Microbiol. (2023) 14:1273364.
40. Donnellan S, Tran L, Johnston H, McLuckie J, Stevenson K, Stone V. A rapid screening assay for identifying mycobacteria targeted nanoparticle antibiotics. Nanotoxicology. (2016) 10(6):761–9.
41. Natan M, Banin E. From nano to micro: using nanotechnology to combat microorganisms and their multidrug resistance. FEMS Microbiol Rev. (2017) 41(3):302–22.
42. Docter D, Westmeier D, Markiewicz M, Stolte S, Knauer SK, Stauber RH. The nanoparticle biomolecule corona: lessons learned–challenge accepted? Chem Soc Rev. (2015) 44(17):6094–121.
43. Horev B, Klein MI, Hwang G, Li Y, Kim D, Koo H , et al. pH-activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano. (2015) 9(3):2390–404.
44. Abed N, Couvreur P. Nanocarriers for antibiotics: a promising solution to treat intracellular bacterial infections. Int J Antimicrob Agents. (2014) 43(6):485–96.
45. Pfaller MA, Diekema D. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. (2007) 20(1):133–63.
46. Kim KJ, Sung WS, Moon SK, Choi JS, Kim JG, Lee DG. Antifungal effect of silver nanoparticles on dermatophytes. J Microbiol Biotechnol. (2008) 18(8):1482–4.
47. Singh J, Vishwakarma K, Ramawat N, Rai P, Singh VK, Mishra RK , et al. Nanomaterials and microbes’ interactions: a contemporary overview. 3 Biotech. (2019) 9(3):68.
48. Ayati MH, Araj-Khodaei M, Haghgouei T, Ahmadalipour A, Mobed A, Sanaie S. Biosensors: the nanomaterial-based method in detection of human gut microbiota. Mater Chem Phys. (2023) 307:127854.
49. Tang J, Ouyang Q, Li Y, Zhang P, Jin W, Qu S , et al. Nanomaterials for delivering antibiotics in the therapy of pneumonia. Int J Mol Sci. (2022) 23(24):15738.
50. Pandey T, Pandey V. Microbial assistance in nano-carrier development: Innovative strategies in drug delivery. J Drug Deliv Sci Technol. (2024) 95:105607.
51. Sohm B, Immel F, Bauda P, Pagnout C. Insight into the primary mode of action of TiO2 nanoparticles on Escherichia coli in the dark. Proteomics. (2015) 15(1):98–113.
52. Burke DJ, Pietrasiak N, Situ SF, Abenojar EC, Porche M, Kraj P , et al. Iron oxide and titanium dioxide nanoparticle effects on plant performance and root associated microbes. Int J Mol Sci. (2015) 16(10):23 630–50.
53. Li J, Yang S, Lei R, Gu W, Qin Y, Ma S , et al. Oral administration of rutile and anatase TiO2 nanoparticles shifts mouse gut microbiota structure. Nanoscale. (2018) 10(16):7736–45.
54. Williams K, Milner J, Boudreau MD, Gokulan K, Cerniglia CE, Khare S. Effects of subchronic exposure of silver nanoparticles on intestinal microbiota and gut-associated immune responses in the ileum of Sprague-Dawley rats. Nanotoxicology. (2015) 9(3):279–89.
55. Jones N, Ray B, Ranjit KT, Manna AC. Antibacterial activity of ZnO nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol Letters. (2008) 279(1):71–6.
56. Banik BL, Fattahi P, Brown JL. Polymeric nanoparticles: the future of nanomedicine. Wiley Interdiscip Rev. (2016) 8(2):271–99.
57. Karimi M, Ghasemi A, Zangabad PS, Rahighi R, Basri SMM, Mirshekari H , et al. Smart micro/nanoparticles in stimulus-responsive drug/gene delivery systems. Chem Soc Rev. (2016) 45(5):1457–501.
58. Bakshi HA, Quinn GA, Aljabali AA, Hakkim FL, Farzand R, Nasef MM , et al. Exploiting the metabolism of the gut microbiome as a vehicle for targeted drug delivery to the colon. Pharmaceuticals. (2021) 14(12):1211.
59. Wojciechowska O, Costabile A, Kujawska M. The gut microbiome meets nanomaterials: exposure and interplay with graphene nanoparticles. Nanoscale Adv. (2023) 5(23):6349–64.
60. Lee BC, Lee JY, Kim J, Shin N, Yoo JM, Kang I , et al. Oral administration of microbiome-friendly graphene quantum dots as therapy for colitis. 2D Mater. (2021) 8(2):025036.
61. Han J, Zhao D, Li D, Wang X, Jin Z, Zhao K. Polymer-based nanomaterials and applications for vaccines and drugs. Polymers. (2018) 10(1):31.
62. Li L, Lin SL, Deng L, Liu ZG. Potential use of chitosan nanoparticles for oral delivery of DNA vaccine in black seabream Acanthopagrus schlegelii Bleeker to protect from Vibrio parahaemolyticus. J Fish Dis. (2013) 36(12):987–95.
63. Illum L. Nanoparticulate systems for nasal delivery of drugs: a real improvement over simple systems? J Pharm Sci. (2007) 96(3):473–83.
64. Yan X, Sha X. Nanoparticle-mediated strategies for enhanced drug penetration and retention in the airway mucosa. Pharmaceutics. (2023) 15(10):2457.
65. Yahya EB, Abdulsamad MA, Aburowais AS, Abogmaza AF. Insights into microbiome-based therapeutics: engineered probiotics, bacteriophages, and microbiome-derived metabolites. Biomed Res Ther. (2025) 12(12):8062–71.
66. Sahu BK, Pradhan L. Nanoparticle-induced stress in environmental microbiomes: ecotoxicological perspectives. Int J Sci Res Eng Trends. (2025).
67. Khan IA, Yu T, Yang M, Liu J, Chen Z. A systematic review of toxicity, biodistribution, and biosafety in upconversion nanomaterials: critical insights into toxicity mitigation strategies and future directions for safe applications. BME Front. (2025) 6:0120.
68. Kumar R, Dutt S, Tripathi AD, Singh AK, Chaturvedi VK, Singh SK. Navigating safety and toxicity challenges in nanomedicine: strategies, assessment, and mitigation. In: Kumar R, Dutt S editors. Nanomedicine: Innovations, Applications, and Breakthroughs in the Quest for Health and Medicine’s Future. Cham: Springer Nature Switzerland (2024). p. 15–37.
69. Bi Y, Xie S, Li Z, Dong S, Teng L. Precise nanoscale fabrication technologies, the “last mile” of medicinal development. Acta Pharmaceutica Sinica B. (2025) 15(5):2372–401.
70. Abdussalam-Mohammed WJJOCR. Review of therapeutic applications of nanotechnology in medicine field and its side effects. J Chem Rev. (2019) 1(3):243–51.
71. Shanmugam H, Rengarajan C, Nataraj S, Sharma A. Interactions of plant food bioactives-loaded nano delivery systems at the nano-bio interface and its pharmacokinetics: an overview. Food Front. (2022) 3(2):256–75.
72. Desai N, Rana D, Patel M, Bajwa N, Prasad R, Vora LK. Nanoparticle therapeutics in clinical perspective: classification, marketed products, and regulatory landscape. Small. (2025) 21(29):2502315.
73. Strain R, Stanton C, Ross RP. Effect of diet on pathogen performance in the microbiome. Microbiome research reports. (2022) 1(2):13.
© The Author(s). 2026 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.